Contorno rosso: guancia estesamente gialla in P. dominula, per lo più nera in P. gallicus;
Freccia blu: mandibola, nera o con poco giallo in P. dominula, estesamente gialla in P. gallicus;
Freccia verde: ultimo sternite giallo in P. dominula, totalmente o prevalentemente nero in P. gallicus.
Freccia bianca: urosterno estesamente nero in P. nimpha; variabile ma sempre con apice bruno in P. associus.
Ocelli equidistanti in P. nimpha; ocelli posteriori più vicini che a quello anteriore in P. associus.
MASCHI
Flagello antennale dorsalmente annerito con ultimo segmento sottile e allungato in P. nimpha e P. associus; flagello antennale dorsalmente chiaro con ultimo segmento non particolarmente allungato in P. gallicus e P. dominula (possibili esemplari ambigui).
Clipeo squadrato con margine inferiore angolato in P. nimpha; clipeo convergente verso il basso e con margine inferiore arrotondato in P. associus (prego notare le due creste ai margini laterali del clipeo).
Clipeo squadrato e con lievi creste laterali in P. nimpha; clipeo squadrato ma piatto in P. dominula; clipeo convergente con definite pieghe laterali in P. associus; clipeo convergente ma piatto in P. gallicus.
Oggi vi parlo di un argomento che volevo affrontare da tempo. Molti fra voi sapranno che gli imenotteri sono il gruppo di animali che mi interessa ed appassiona maggiormente, ma un’altra mia passione è la biologia evolutiva. Quale miglior modo per unire questi miei due interessi se non parlando del singolare percorso evolutivo degli imenotteri? Ne sono personalmente affascinato, non solo perché tale strada è molto ricca di “colpi di scena” e adattamenti pazzeschi, ma anche perché offe numerosi esempi di fenomeni evolutivi poco discussi ma affascinanti come ad esempio l’exaptation, di cui parleremo in seguito nel dettaglio.
Il nostro percorso inizia nel Permiano, circa 280 milioni di anni fa. All’epoca, l’antenato comune di tutti gli imenotteri era un esserino dall’aspetto anonimo le cui larve, provviste di zampe, pseudozampe e una buona locomozione, potevano tranquillamente passare per bruchi. Tali larve si cibavano di materiale vegetale, primariamente getti e germogli verdi, che sgranocchiavano dall’esterno o nei quali scavavano piccole gallerie. Utilizzando l’odierna classificazione tassonomica, tale esserino era un Synphyta (sinfite), ovvero un imenottero con torace e addome fusi assieme a formare un blocco corporeo pressoché unico, con cuticola morbida e al termine dell’addome una formazione tozza e corta ma anche molto rigida, che l’organismo utilizza per incidere gli steli delle piante e deporre le uova in queste incisioni.
Xyelidae non altrimenti specificato (fonte: iNaturalist), questo è probabilmente l’aspetto approssimativo che doveva avere l’imenottero ancestrale
Inizialmente non vi fu molta diversificazione, ma dopo la più grande estinzione di massa della storia della Terra, che aprì molte nicchie ecologiche nuove ma segnò anche la fine del Permiano e l’inizio del periodo Triassico, avvenne una rapida radiazione adattiva che portò all’evoluzione di numerosi gruppi con specializzazioni diverse. La radiazione adattiva è quel processo che consente l’evoluzione di numerose specie in un lasso di tempo relativamente breve (pochi milioni di anni). Tale fenomeno è spesso innescato dall’apertura di una grossa nicchia ecologica tutta da colmare, come accade spesso proprio in seguito ad un’estinzione di massa. In alcuni gruppi che si originarono a seguito di questa radiazione, vi erano specie le cui larve non si nutrivano più di getti verdi delle piante, ma dello stesso legno di tronchi e rami: ad esempio il gruppo noto come Siricoidei. a questo punto le larve degli imenotteri iniziano ad abbozzarsi, perdono le zampe e le pseudozampe, perdono buona parte della loro mobilità.
Siricidae del genere Tremex mentre perfora un pezzo di legno per deporvi le uova
La vita di una larva di Siricoideo è abbastanza frugale ma onesta: nasci, ti scavi una galleria nel legno, mangi legno la mattina, mangi legno la sera, fai una galleria qua, fai una galleria là, ogni tanto fai la cacca, poi quando sei pronto ti crei un bozzolo formato dalle tue stesse feci nel quale affronti la fase pupale, e dopo un po’ di tempo ti trasformi in un esemplare adulto e svolazzante pronto a usare un organo stretto e rigido, l’ovopositore, per trovare altro legno nel quale trapanare per deporre le uova. Ricordiamo però, che nell’evoluzione va avanti chi riesce ad adattarsi meglio all’ambiente.
Adattarsi significa anche riuscire a nutrirsi adeguatamente, e il legno non è che sia proprio la colazione del campione… la cellulosa e la lignina sono difficilissime da digerire, serve per forza fare simbiosi con delle colonie di microrganismi che poi ti vivono nell’intestino, e la resa energetica non è mica altissima… nsomma, mangi per campare. Dunque potreste mai biasimare una larvetta di Siricide che fra un boccone al legno e l’altro, ogni tanto, giusto per cambiare un po’, tira un mozzico ad un’altra larva che le capita a tiro? ma sì, è un’indulgenza sporadica, una tantum, dai, chiudiamo un occhio per questa volta, oggi mangio mio cugino e domani torno a mangiare legno, che vuoi che succeda…
Beh succede che nasce una categoria di Sinfiti, gli Orussidae, le cui larve ben presto imparano a nutrirsi solo di larve di altri sinfitiMamma Orusside cerca una larva bella succulenta, usando le tracce chimiche lasciate dalle feci di tale larva disseminate in giro. Una volta trovata, vi depone un ovetto vicino, o magari proprio sopra, o perché no, anche dentro, tanto l’ovopositore se sai come manovrarlo può fungere un po’ anche da ago ipodermico. Quando la larva dell’Orusside emerge, essa si trova già un banchetto riccamente proteico li a portata di mano e divora la larva ospite. Signore e signori, ecco che nasce il parassitoidismo!
Orussus terminalis, famiglia Orussidae, ovvero gli inventori del parassitoidismo negli imenotteri. Credit iNaturalist
R I V O L U Z I O N E. La strategia di mamma Orusside riscuote un successo enorme, a quanto pare parassitare un’altra specie è molto più vantaggioso in termini calorici che mangiare legno, chi l’avrebbe mai detto! Ma aspetta un momento… la fuori ci sono anche un sacco di altri insetti con larve succose e ricche di proteine, ci sono i coleotteri, ci sono i lepidotteri… sta a vedere che posso parassitare anche loro?? E così, altra enorme nicchia ecologica da colmare, altra radiazione adattiva, si evolve una quantità scandalosa di nuove specie di parassitoidi.
Sembra proprio che questa strategia riproduttiva sia perfetta, così perfetta che emergono addirittura parassitoidi degli stessi Orussidi! Il cugino di primo grado dell’Orusside, tale signor Apocrito, che è anch’esso un parassitoide ma non è mica scemo, evolve un modo per far sì che le sue larve non vengano rintracciate da altri parassitoidi: la stipsino, davvero: dal momento della nascita fino a poco prima di impuparsi, la larva dell’Imenottero Apocrito è priva di ano. Mantiene tutti gli scarti digestivi dentro il corpo, per evitare che l’aroma delle sue feci attiri cugini indesiderati.
Trogus lapidator, Ichneumonidae, parassitoide del bruco di macaone
Gli Apocrita sono dei rivoluzionari, sviluppano anche un’interessante novità: al fine di manovrare meglio l’ovopositore -anche perché inoculare uova in un ospite semovente è un lavoro certosino che richiede una precisione più che chirurgica- sviluppano una strozzatura nel mezzo del corpo, fra il primo e il secondo segmento addominale. Il primo segmento rimane fuso col torace, nella parte anatomica detta propodeo, mentre fra quest’ultimo e il resto dell’addome si genera un “vitino” ristretto chiamato peziolo. Apocrita significa proprio questo, dal Greco Apokriton che significa separato. Il peziolo non solo rende più facile l’inoculazione di uova all’interno dell’ospite, ma stabilizza anche il volo. Infatti a che alcuni Ditteri, abilissimi volatori, l’hanno evoluto indipendentemente. Alcuni ditteri hanno anche evoluto il parassitoidismo. Insomma, sono un po’ copioni dai.
Bene, ora che la radiazione evolutiva dei parassitoidi è compiuta, e gli imenotteri hanno trovato il modo perfetto per vivere, ovvero a spese di altre specie, ci si può godere in pace il resto del Mesozoico: giusto in tempo per guardare l’ascesa e il dominio dei dinosauri, che fino ad ora erano ancora insulsi prototipi!
E invece no! Perché l’evoluzione non si ferma mai! Non esiste la perfezione, la vita è cambiamento. Alcuni Apocrita, precisamente nel gruppo Aculeata, caratterizzato da un ovopositore retrattile, hanno notato che per aumentare le chances di successo della loro prole si può costruire un nido.. ma nulla di troppo elaborato, che so, una celletta di fango, un buco nel terreno… e murarci vive le prede in compagnia delle proprie uova. Tecnicamente non si parla più di parassitoidismo, ma di una forma un po’ contorta di predazione. Ed è qui che troviamo l’exaptation: dal momento che non è più necessario inoculare l’uovo nell’insetto ospite, perché tanto chiuso nel nido mica scappa, non serve più avere un ago che sporge dalle parti basse. L’ovopositore perde la funzione per la quale la natura lo ha disegnato. E quindi che fa, sparisce? eh no! perché è molto più facile murare viva una preda se questa è anestetizzata… Si insomma, collabora meglio. Capite già dove vado a parare? è così che l’ovopositore diventa un mezzo di inoculazione non delle uova, ma del veleno. Un veleno blando, che serve solo a tramortire le prede, nulla di troppo elaborato per ora… ma sempre veleno è!
Sceliphron destillatorium, Sphecidae, predatore solitario di ragni che accumula in nidi fangos
Ridendo e scherzando, anzi, ridendo e predando, siamo già nel bel mezzo del Giurassico (200-150 milioni d’anni fa). Alcuni Apocrita predatori hanno notato che ultimamente alcune delle loro prede sono sporche di questa strana robetta gialla… com’è che si chiama, polline? bleah che schifo, sa di polvere, meglio dare una lustrata alle prede prima di metterle nel nido. Dicevamo? ah sì, i predatori col nido. Inutile girarci intorno, l’unione fa la forza, così ben presto alcune specie di predatori solitari inventano forme primitive di cooperazione. Hanno questa strana idea in mente che se parte della prole, invece che andar via a formarsi il proprio nido, resta ad aiutare la mamma a crescere altra prole, le possibilità di successo della famiglia aumentano. Avrà fortuna questa loro strampalata idea? boh, staremo a vedere.
E siamo nel Cretaceo (150-66 milioni d’anni fa). L’era di picco dei dinosauri più noti e fighi. Le angiosperme stanno diventando predominanti, fiori ovunque, il polline diventa una presenza sempre più forte nella vita degli imenotteri predatori. La signora Anthophila, che essendo moderna e di larghe vedute non condivide il pregiudizio delle sue sorelle nei confronti di questa nuova sostanza, si è adattata ad usare prede ricoperte di polline. Un bel giorno si è accorta che il polline è altrettanto nutriente e proteico, ed è molto molto abbondante, anche perché non bisogna cacciare per ottenerlo, le piante lo danno gratis. Così fa una cosa pazza e assurda, abbandona la dieta carnivora e inizia a nutrire le sue larve con solo polline. Mah dai, non è così male. Sembra che la fortuna sia dalla sua part
BOOOOOOOM
asteroide, ciao ciao dinosauri. E mo’? è la fine? No. I fiori sono sopravvissuti all’estinzione, anzi si stanno diversificando un sacco in questo nuovo mondo post-apocalittico. Radiazione evolutiva delle api, tante nuove specie che si specializzano in vari modi per raccogliere il polline, alcune sulle zampe posteriori, altre sulla pancia, certe addirittura non contente si mettono a raccogliere anche il nettare. Per ora sono tutte solitarie, fin quando ad una di esse viene la brillante idea di mettere su famiglia. ricordate le vespette predatrici di prima, che volevano creare una società? beh l’idea non è proprio brutta, e il sistema aplodiploide di determinazione del sesso, condiviso da tutti gli imenotteri e che ha la peculiare conseguenza di rendere ogni femmina più genticamente imparentata alle proprie sorelle rispetto che alla propria madre, sembra agevolare la comparsa di una cooperazione eusociale: regina riproduttrice; operaie sterili, sorelle che collaborano fra loro alla crescita di altre sorelle; maschietti sporadici giusto per mescolare i geni ogni tanto. L’idea ha un successo tale che avviene un’altra mini radiazione evolutiva nel corso del Cenozoioco, l’era corrente, che porta alla nascita di gruppi interi di api eusociali quali i Meliponini, gli Euglossini, i bombi e in fine l’ape mellifera. Il materiale di nidificazione non è più l’obsoleto fango, ma la versatile cera prodotta da ghiandole sul loro stesso corpo.
Bombus ruderatus, Anthophila, eusociale avanzato con una divisione in caste netta
Nel frattempo anche le vespe di prima hanno intrapreso una strada simile, hanno scoperto che l’unione fa davvero la forza, e che il super-organismo può fare cose che sono impossibili per il singolo individuo. Tant’è che fra Cretaceo e Paleogene l’eusocialità, oltre che nelle api, si è evoluta ben tre volte: prima nei Formicidae, anche dette formiche, la cui casta operaia ha perso le ali. Poi altre due volte nei Vespidae.
Polistes dominula, Vespide eusociale primitivo con una divisione in caste ancora non netta
Vogliamo parlare dei Vespidae? beh che dire. Innanzitutto, come appena accennato, sono arrivati all’eusocialità due volte indipendentemente: la prima volta negli Stenogastrini, un gruppo limitato al Sud Est Asiatico, la seconda volta nell’antenato comune a Vespini e Polistini, fra cui annoveriamo i famigerati calabroni e le vespe “classiche” che noi tutti conosciamo. Ma non solo! alcuni di essi, i Masarinae, sono arrivati alla stessa conclusione degli Anthophila: hanno abbandonato lo stile di vita da cacciatori in favore ad una dieta basata sul polline, e ora vanno in giro a farsi chiamare “Pollen wasps”
Pseudomasaris sp, un Masarinae, tiene a farvi sapere che gli Anthophila se la tirano troppo e che anche lui ha imparato ad usare il polline come alimento larvale, ma resta umile (iNaturalist)
Ma volete sapere qual è la parte sorprendente di tutto ciò? per ricostruire la storia evolutiva degli imenotteri non abbiamo bisogno di andare a cercare fossili: abbiamo ogni passo del loro percorso nel nostro giardino, ancora vivo e vegeto. esistono ancora tanti sinfiti che mangiano le piante. Esistono ancora tanti (ma proprio taaaaanti) parassitoidi che inoculano uova nelle loro vittime. Esistono ancora un sacco di predatori solitari, o di specie presociali.
Non voglio che passi il messaggio che l’evoluzione di questi organismi sia stata una scala a progressi dove ogni tappa è stata un miglioramento della precedente in termini assoluti. Tutte le strategie di vita che ho descritto, dal sinfita sempliciotto del Permiano fino ai Masarinae odierni, sono strategie vincenti ancora oggi adottate da gruppi che possiamo osservare facilmente. Allego una carrellata di foto delle guest stars di oggi, a partire dai gruppi con stile di vita più “ancestrale” per finire con quelli più “sofisticati”.
Grazie per il vostro tempo e per essere arrivati in fondo a questo gigantesco post
Visto l’interesse recente che si è creato nei confronti di Vespa orientalis, ho deciso di spiegare tutto ciò che c’è da sapere a grandi linee in un unico post.
Si, è vero che una colonia di V. orientalis è stata di recente (settembre 2021) rinvenuta a Grosseto (fra l’altro da parte di un membro di questo gruppo). No, non è vero che è una specie aliena arrivata dall’oriente per invaderci tutti e bullizzare le nostre apine. Cerco di fare un po’ di chiarezza punto per punto:
Vespa orientalis a Sliema, Malta 2018
DISTRIBUZIONE NATURALE:
L’epiteto specifico “orientalis” sembra proprio implicare che sia l’ennesimo insetto venuto a invaderci dall’Asia, ma ciò non è assolutamente vero. Il nome della specie allude NON alla sua provenienza, ma alla località nella quale sono stati raccolti gli esemplari-tipo, ovvero gli esemplari sui quali Carl Von Linné (per gli amici Linneo) si basò per descrivere la specie nel 1771. “Ex Oriente” è da intendersi come una non meglio precisata località a est del Mediterraneo.
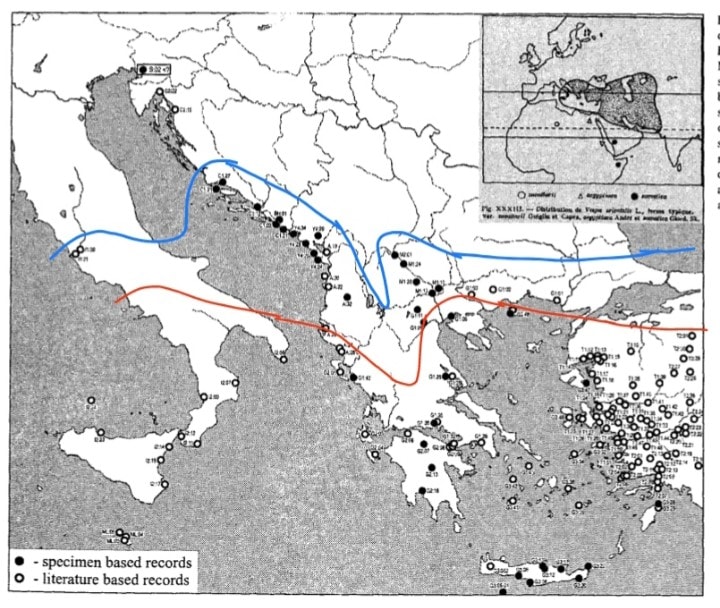
La sua distribuzione è molto ampia, include l’Asia sudoccidentale, da Turchia e Arabia fino all’India; l’Asia centrale a nord fino al Kazakistan; l’Africa settentrionale con numerose popolazioni nel Sahara e giù fino al corno d’Africa; la Penisola balcanica Meridionale inclusa Grecia e vari arcipelaghi annessi; Cipro; Creta; Malta; l’Italia Meridionale dalla Campania in giù compresa la Sicilia. La specie è SEMPRE stata presente nelle zone che ho appena elencato.
Ritengo interessante, a fine di completezza, menzionare che secondo alcuni è plausibile che sia stata portata da noi tramite commercio navale in epoca Fenicia, circa 3000 anni fa, ma la tesi preponderante è che sia un specie trans-adriatica, giunta da noi in modo naturale in epoca preistorica, probabilmente durante uno dei tanti episodi di prosciugamento del Mar Adriatico. In ogni caso è presente in Sud Italia almeno da millenni, per cui è da considerarsi autoctona in ogni senso del termine.
Il range periferico di Vespa orientalis in Europa è soggetto a continue espansioni e ritiri, in quanto questa specie è legata a climi caldi e almeno in parte aridi. Infatti, nella seconda metà degli anni ’60, un trend di raffreddamento climatico che ha colpito l’Europa meridionale ha causato la completa sparizione di Vespa orientalis da città come Roma, Split e Skopje, tutte aree nelle quali era precedentemente segnalata. Nello stesso periodo persino a Malta, pur non essendo sparita, la specie pare aver subito una drastica rarefazione.
Cetkovic (2004) nota che il requisito principale affinché una determinata località sia in grado di ospitare Vespa orientalis è la media termica del mese più caldo sopra i 25.5°. In parole povere, se la temperatura media giorno/notte di una determinata località è sopra questa soglia per almeno un mese, Vespa orientalis è almeno teoricamente capace di viverci.
Le aree italiane con questo requisito sono le coste sicule, quelle calabresi, la pianura pugliese e campana: tutte zone nelle quali effettivamente Vespa orientalis è presente e abbondante da sempre. Se invece si va verso nord, si arriva a un punto, fra Campania e Lazio, nel quale la soglia dei 25.5°C estivi non è raggiunta, difatti storicamente il suo territorio si ferma proprio lì.
Allora come mai ci sono sempre più segnalazioni da aree come Roma, Grosseto e addirittura Trieste? Semplice. Queste sono tutte aree urbane soggette all’effetto isola di calore, che tende a rendere qualsiasi città di dimensioni significative alcuni gradi più calda rispetto alla campagna circostante. L’espansione delle aree urbane ha facilitato la risalita di Vespa orientalis lungo lo stivale, anche aiutata dal trasporto accidentale umano, tramite il quale essa “salta” da un’isola di calore all’altra.
Dettaglio del capo di Vespa orientalis
COSA ASPETTARSI:
Sicuramente in futuro sempre più città del centro-nord verranno raggiunte da questa specie, creando dal Lazio in su una distribuzione a macchia di leopardo che salta le aree più rurali. Dubito personalmente che arriverà a colonizzare l’area Padana, perché troppo umida, o le aree montane per ovvi motivi.
La pressione predatoria nei confronti delle api al centro-nord sarà quindi massima nei pressi delle zone urbane e minima nelle campagne, questo almeno finché il riscaldamento globale non porterà anche la campagna oltre la menzionata soglia dei 25.5°C, cosa che potrebbe senz’altro accadere ma fra vari decenni.
Un esemplare di V. orientalis dalla mia collezione (Uzbekistan, 2014)
È vero che in densità sufficientemente alte Vespa orientalis può imprimere alle api da miele una pressione predatoria molto intensa. Ma in base ai (pochi) dati attualmente a disposizione, pare che questo sia per ora un problema limitato alle isole di calore di Sicilia e Campania. Non è probabile che ciò avvenga anche al centro-nord nelle condizioni climatiche attuali.
Ah, e per favore NON CONFONDIAMO V. orientalis con V. velutina, sono due specie differenti con storie, distribuzioni ed ecologie differenti. Quest’ultima è per ora presente solo nelle regioni del Nord Ovest (Liguria, sud del Piemonte, costa settentrionale della Toscana), e ha una pressione predatoria nettamente superiore nei confronti delle api da miele a parità di densità.
Ultimamente mi sono interessato al curioso caso delle api del Capo sudafricane, e mi piacerebbe condividerla con voi, sia per condensare le informazioni che ho raggruppato fino ad ora che per rendervi partecipi di questo caso eccezionale della biologia degli imenotteri. Buona lettura!
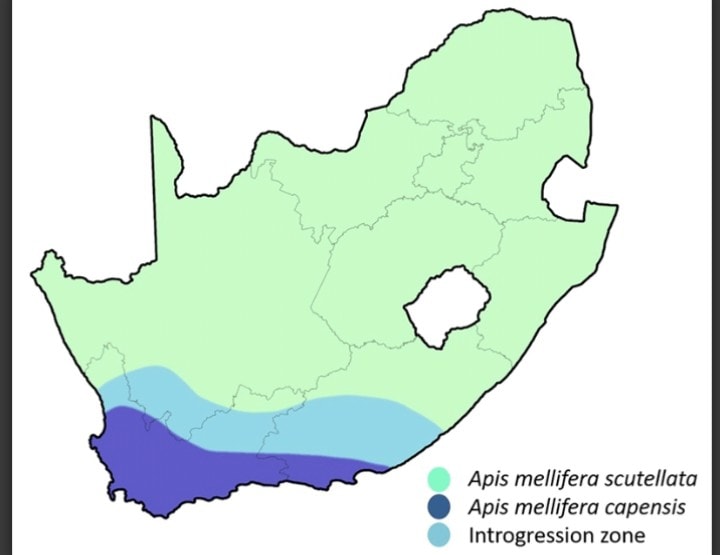
Apis mellifera, “l’ape da miele occidentale”, è una specie di interesse economico e agricolo per l’essere umano allevata da millenni. È volgarmente detta “ape Europea”, in quanto i ceppi più allevati in tutto il mondo hanno origine in Europa (prima fra tutte la sottospecie “ligustica” di origine italiana), tuttavia la specie copre un areale molto più vasto dell’Europa, spingendosi fino all’Asia centrale e all’intero Continente Africano. Attraverso questo areale, essa si frammenta in varie sottospecie, ognuna tipica di una determinata località.Il Sud Africa ospita due di queste sottospecie: una è Apis mellifera scutellata, la più comune e diffusa, suonerà familiare a qualcuno di voi in quanto è uno dei progenitori delle così dette “api assassine” o “africanizzate”, è diffusa in quasi tutto il S.Africa e in buona parte dell’Africa orientale. La seconda sottospecie, Apis mellifera capensis, è invece endemica della regione del Capo, nell’estremo sud del Paese, e la sua distribuzione ricalca quella del bioma noto come “Fynbos”, ovvero la versione Sudafricana della macchia mediterranea (infatti la regione del Capo è caratterizzata da un clima mediterraneo, con inverni freschi e umidi ed estati siccitose). Esiste anche una fascia di sovrapposizione a nord del Capo nella quale entrambe le sottospecie sono presenti e si ibridano regolarmente.
IL SUPERPOTERE DELL’APE DEL CAPO
Tutti siamo familiari con la struttura di una colonia di api da miele: esiste una regina, in grado di deporre le uova, migliaia di operaie normalmente sterili che difendono la colonia, nutrono la covata e procacciano nettare e polline, e qualche manciata di maschi detti fuchi che hanno il compito di fecondare una regina giovane, in modo che essa ne utilizzi gli spermatozoi per fecondare le uova che deporrà lungo l’arco della sua vita. La fecondazione, che avviene in un volo nuziale, è necessaria per produrre femmine (sia operaie che regine), mentre la deposizione di un uovo non fecondato risulterà in un maschio.
In una colonia di api da miele “comuni”, quando una regina muore o sparisce, le operaie della colonia rimasta orfana selezionano alcune giovani larve femminili e le utilizzano per ottenere regine d’emergenza. Se la precedente regina sparisce senza lasciare uova, e quindi la colonia non ha larve femmina abbastanza giovani da convertire in regine, alcune operaie inizieranno a deporre le uova, ma non essendo fecondate potranno solo generare fuchi. Senza più rinnovo generazionale, la colonia si estinguerà in poche settimane. Queste colonie sono dette fucaiole e sono una grossa rottura di scatole per gli apicoltori.
Il bioma del Fynbos è battuto da forti venti primaverili. Questi venti disturbano i voli nuziali delle regine e talvolta le uccidono, rendendo altissimo il tasso di orfanità delle colonie di api del Capo. L’evoluzione, che vede e provvede, ha dunque donato ad alcune operaie di ape del Capo la capacità di clonare sé stesse all’occorrenza, deponendo uova femminili senza il bisogno di accoppiarsi. Questa capacità permette loro di disporre sempre di nuove larvette femminili con le quali sostituire la regina in caso di scomparsa. In questo modo, se la regina sparisce senza lasciare uova, la colonia non diventa fucaiola, semplicemente nuove regine verranno ottenute dalle uova clonali deposte dalle operaie, ripristinando l’ordine gerarchico nella colonia. Questa notevole capacità, chiamata partenogenesi telitoca, fu scoperta nel 1912.
BYPASSARE IL SISTEMA
Il superpotere che hanno queste operaie di capensis (non lo hanno tutte in una colonia, ma solo alcune dette clonali) permette alla sottospecie di prosperare nel Fynbos, ma ha un effetto collaterale: un’operaia capace di deporre uova femminili è anche capace di clonare sé stessa se da queste uova nascono altre operaie con la sua stessa capacità. In questo modo è possibile bypassare il normale sistema eusociale e creare una colonia di sole operaie che generano altre operaie, senza il bisogno di maschi né regine. Nel Fynbos questa cosa non succede, perché essendo le operaie clonali meno fertili delle regine, alla colonia conviene tenersi il sistema “tradizionale” con regina e fuchi e riservare la partenogenesi telitoca ai casi di reale necessità. Ma cosa succede quando si trasporta un’operaia clonale in un’area dove non ci sono colonie di api del Capo? Negli anni ’70 si teorizzò che se una cosa del genere dovesse accadere, essa si introdurrebbe in una colonia di api di un’altra varietà e si comporterebbe da parassita, generando cloni di sé stessa che potrebbero portare all’estinzione della colonia ospite. Negli anni ’90 la teoria divenne pratica, quando degli apicoltori introdussero l’ape del Capo nella provincia del Gauteng, fuori dal loro areale naturale e dentro quello dell’ape scutellata.
LA PIAGA DELLE CAPENSIS
Nel 1990, una singola operaia clonale di ape del Capo si è ritrovata spaesata nel Gauteng, a centinaia di km dal suo amato Fynbos. Si è introdotta in una colonia di ape scutellata, la sottospecie locale, e ha fatto quello che sapeva fare meglio: ha prodotto uova che si sono sviluppate in operaie cloni, che hanno poi fatto la stessa cosa. Nel giro di alcune settimane, la colonia di scutellata si è ritrovata migliaia di capensis che invece di foraggiare deponevano uova tutto il tempo, diminuendo l’importo di polline e nettare, e causando per di più la morte della regina di scutellata, che le capensis vedono come una competitrice riproduttiva. Col passare del tempo la colonia ospite collassa sotto il peso di tutte quelle cloni di capensis da mantenere, estinguendosi completamente nel giro di qualche mese. A quel punto le capensis abbandonano la colonia ormai distrutta e ne cercano altre, ripetendo lo stesso copione, insediandosi e riproducendosi come dei virus. Nel 1991, cloni di cloni di cloni di cloni di cloni di quella singola operaia hanno portato alla distruzione, nel Gauteng e zone limitrofe, di oltre 30.000 colonie di scutellata. Il fenomeno è stato ribattezzato la piaga delle capensis, e severissime contromisure sono state adottate per limitarne i danni, come alveari-trappola e severissime leggi.
La distribuzione delle sottospecie in Sud Africa, compresa la zona di ibridazione fra le due.
Questa linea genetica di cloni di cloni di cloni di cloni di cloni di cloni di cloni di una singola operaia iniziale continua ancora oggi, dopo 30 anni, ed è fra le principali avversità dell’apicoltura del Gauteng e regioni circostanti. Per legge, oggi, se un apicoltore si rende conto di avere le capensis in una delle sue famiglie di scutellata, è tenuto a distruggerla per evitare che esse si diffondano.
La distruzione non è facile: se la regina delle scutellata è ancora presente, si può provare a separare le scutellata in una nuova arnia e bruciare quella vecchia per distruggere la covata di capensis. Se invece la regina è già andata e la colonia è invasa dalle capensis, l’unica è chiamare un disinfestatore per debellare l’intera colonia con insetticidi.
La cosa è resa ancora più difficoltosa dal fatto che appena si apre un’arnia invasa dalle capensis, alcune di esse tenderanno a scappare e trovare rifugio nelle arnie vicine, infettandole a loro volta. Il problema è limitato al Sud Africa, ma se la capensis fosse introdotta in Europa o in altre località, potrebbe diventare una seria minaccia all’apicoltura locale. Fra l’altro sarebbe perfettamente in grado di sopravvivere nel clima Italiano, che è pressoché uguale a quello del Capo a parte per le stagioni sfasate, per cui l’unica è sperare che le capensis non vengano mai Introdotte qui. Basta una sola operaia clonale per generare un’intestazione che dura decenni.
Le operaie di capensis sono più scure di quelle di scutellata, hanno un addome tendente al nero, per cui in genere sono facili da riconoscere, come mostra la prima foto, ma non è sempre una differenza così ovvia. La loro colorazione scura è simile a quella di alcuni ceppi Europei, ciò significa che se venisse Introdotta qui probabilmente ci metteremmo un po’ ad accorgercene. Nella seconda foto la distribuzione sudafricana delle due sottospecie e la zona di overlap.
Il ciclo vitale dei calabroni (genere Vespa) si compone generalmente di sei fasi:
FASE SOLITARIAanche detta fondazione: è la fase nella quale la regina, in primavera, si risveglia dal torpore invernale e inizia la costruzione del nido. Essa crea le prime cellette, un abbozzo di involucro, depone le prime uova e alleva la prima nidiata di larve. In questa fase la regina si occupa da sola di tutto: caccia, difesa, costruzione, cura della covata. Questa fase termina con la nascita delle prime operaie.
FASE COOPERATIVA: è la fase nella quale le prime operaie emergono e iniziano a svolgere i loro compiti, aiutando la regina nella caccia e nella costruzione.
FASE POLIETICA: inizia quando la regina cessa ogni attività fuori dal nido e si dedica esclusivamente alla deposizione delle uova, lasciando alle operaie, in costante crescita numerica, tutti i lavori pesanti. Questa è la fase che vede il maggior sviluppo del nido e della colonia in termini di numero di esemplari.
FASE SESSUALE o riproduttiva: inizia alla nascita del primo esemplare fertile (in genere un maschio) e marca la maturità sessuale della colonia. In questa fase, la produzione di operaie inizia a calare e quella di sessuati (maschi e nuove regine) aumenta esponenzialmente. Si raggiunge ora il picco numerico della colonia.
DISSOLUZIONE: in questa fase, anche detta fase abortiva, maschi e nuove regine abbandonano la colonia per accoppiarsi, le ultime larve rimaste vengono abortite, la regina anziana muore e le operaie decrescono rapidamente fino all’estinzione della colonia. I maschi fertili di Vespa mandarinia e Vespa soror nascono qualche settimana prima delle femmine e iniziano ad esplorare il territorio alla ricerca delle altre colonie in modo da potersi accoppiare con le regine di tali colonie appena queste escono dal loro nido. Gli accoppiamenti di Vespa velutina, Vespa orientalis e Vespa crabro avvengono in zone in rilievo come pali, alberi, edifici, ecc.
SVERNAMENTO: in questa fase, le nuove regine dopo essersi accoppiate, si cercano un posto per passare l’inverno. Intanto i maschi muoiono, lasciando le regine quiescenti gli unici esemplari presenti in questa parte del ciclo. Termina con l’inizio di una nuova fase solitaria la primavera seguente, quando il ciclo ricomincia. Si stima che solo il 10% delle regine riesca a fondare una nuova colonia, il restante 90% non sopravvive abbastanza a lungo oppure non riesce ad accoppiarsi con successo (caso frequente soprattutto in Vespa mandarinia e Vespa soror).
Tabella di confronto
Vespa crabro
Vespa orientalis
Vespa velutina
Vespa mandarinia
Quantità di operaie nido primario
qualche decina
–
qualche decina
qualche decina
Dimensione nido primario
–
7 – 8 cm
–
Località nido primario
Posizione riparata
Posizione riparata o cavità
Posizione riparata
Cavità sotterranea o all’interno di tronchi
Vita media operaia
30 giorni
30 giorni
30 giorni
30 giorni
Alimentazione operaie
glicifago
glicifago
glicifago
glicifago
Alimentazione larve
proteine animali
proteine animali
proteine animali
proteine animali
Località nido secondario
Posizione riparata
–
Rami di alberi
–
Dimensione nido secondario
40 – 70 cm
30 – 50 cm
70 cm – 1 metro
50 cm – 1 metro
Numero stimato di nuove regine per colonia
50 – 150
50 – 150
1000 – 1500
200 – 500
Solitaria
Aprile Maggio
Marzo Aprile
Marzo Aprile
Maggio Giugno
Cooperativa
Giugno
Maggio Giugno
Aprile Maggio
Giugno Luglio
Migrazione nido secondario
–
–
Giugno Luglio
–
Polietica
Luglio Agosto
Giugno Luglio
Giugno Agosto
Agosto Settembre
Sessuale
Settembre Ottobre
Agosto Settembre
Settembre Novembre
Settembre Novembre
Dissoluzione
Novembre
Novembre Dicembre
Novembre Gennaio
Novembre Dicembre
Svernamento
Novembre Aprile
Dicembre Marzo
Novembre Febbraio
Novembre Maggio
N. Adulti al picco
300 – 500
300 – 500
2000 – 4000
500 – 1000
Nella prima fila in alto potete vedere gli insetti di cui parliamo. Illustrazione di Daniele Valeriani
Dal momento che è una domanda che viene fatta di frequente, e ancora più frequenti sono le errate identificazioni dei nidi, ho preso in prestito dal web (aprendole vedrete i link rispettivi) due foto secondo me rappresentative di nidi tipici di Vespa velutina e Dolichovespula.
Ormai siamo in inverno, gli alberi caduci hanno quasi tutti perso le foglie, quindi capita molto spesso di trovare queste strutture cartacee appese ai rami. È bene saper distinguere i nidi di queste due vespe, perché una delle due, Vespa velutina, è una specie di origine asiatica che è stata introdotta in Europa per sbaglio. La popolazione europea si è originata in Francia nel 2004 a partire da una o poche regine cinesi giunte molto probabilmente tramite un carico di vasi per bonsai (letteralmente ‘dalla Cina cor furgone’). La presenza di questa specie andrebbe riportata ad enti appositi, in quanto è una specie invasiva che causa danni all’ecosistema nostrano e all’industria apistica. Dolichovespula è invece un genere comprendente alcune specie native dell’Europa che fanno parte del nostro ecosistema da sempre, pertanto non andrebbero distrutte in modo ingiustificato, e non vi è il bisogno di riportarle a nessuno. Vediamo quali sono le differenze nella struttura dei nidi:
FORMA: Il nido di quasi tutte le specie di Dolichovespula, quando appeso ad un ramo o altra superficie, tende ad assumere una tipica forma a pera rovesciata con la parte alta più ampia e quella bassa più stretta. Questa forma accomoda la configurazione dei favi all’Interno: il favo più sviluppato, qualora doveste dissezionare il nido, sarà quasi sempre quello più in alto (il più vecchio). -Al contrario, il nido di V. velutina tende ad avere una forma più globulare o ovoidale, molto spesso con la sommità più stretta rispetto alla parte bassa, probabilmente per far scivolare la pioggia più facilmente. Dissezionandolo, aspettatevi di trovare i favi più larghi verso il basso o verso il centro.
INVOLUCRO: Il nido classico di Dolichovespula tenderà ad avere nella metà più bassa un involucro molto liscio (laminare), cioè i vari strati che lo compongono vanno pochissimo a contatto fra loro, rimanendo paralleli. Nella parte alta del nido, invece, è possibile trovare un più alto grado di anastomosi fra i vari strati, che comporta la formazione di varie piccole sacche. In sezione, troverete che l’involucro è molto più spesso (maggior numero di strati) nella parte bassa, quella laminare, mentre i favi tendono ad essere concentrati in quella alta, che avrà quindi meno strati. -L’involucro di Vespa velutina, invece, è completamente formato da sacche (embricato) e in sezione mostra di avere circa lo stesso spessore sia in basso che in alto.
DIMENSIONI: Ovviamente questo non è un carattere assoluto, dipende dallo stato di avanzamento del nido, ma in genere i nidi di Dolichovespula non sono più grossi di un pallone da calcio. Al loro interno troverete da uno a tre favi, qualche volta fino a cinque, anche se l’ultimo è quasi sempre incompleto. -Le dimensioni del nido di Vespa velutina sono più generose: nidi molto prosperosi possono arrivare a un diametro di quasi un metro, rendendo difficile avvolgere le proprie braccia attorno ad essi, ma in genere si fermano attorno ai 50/70 cm. In sezione mostreranno facilmente una dozzina di favi.
COLORE: Il nido di Dolichovespula è composto da fibre di legno secco, pertanto spesso tende ad assumere un colore grigio chiaro. Non è sempre vero, infatti qualche volta hanno bande di svariati colori a seconda del tipo di legno usato. Ho in collezione un nido con alcune bande rosse che fu costruito in parte usando legno da uno steccato dipinto di rosso. Quasi sempre è possibile distinguere bande di colore bianco, sono i resti degli opercoli delle pupe, che le operaie riciclano. -Il nido di V. velutina è anch’esso a bande, ma si nota di meno, perché il colore è relativamente uniforme e mai grigio. In genere è color ocra, perché ottenuto da legno marcescente o comunque con un certo livello di umidità.
CONSISTENZA: Il nido di Dolichovespula, al tatto, risulta flessibile. Se prendete un pezzo di involucro e lo piegate attorno a una matita, sarà difficile che si spezzi. Al microscopio, le fibre di cellulosa si mostrano strette, lunghe e parallele. -Il nido di V. velutina è invece abbastanza friabile, se sottoposto al test della matita l’involucro tenderà a frammentarsi, e al microscopio le fibre di cellulosa sono irregolari, spezzettate e ricche di impurità.
LOCALITÀ: Dolichovespula è un genere di clima temperato che non ama molto l’ambiente mediterraneo, per cui è più facile incontrarlo in montagna o in aree ombreggiate. La maggioranza delle specie italiane è per lo più alpina, mentre una, Dolichovespula media, è comune lungo gli Appennini, percorrendoli completamente fino alla Calabria. -V. velutina è comune anche a basse altitudini fino al livello del mare, ed è per ora (dicembre 2020) limitata a Liguria, sud del Piemonte e nord-ovest della Toscana. Se non vivete in una di queste zone, per ora, è molto difficile che troviate questa specie.
CICLO VITALE: entrambe le specie hanno colonie annuali che vengono fondate in primavera, si sviluppano in estate e si estinguono con l’arrivo della stagione fredda. Dolichovespula è generalmente attiva dalla tarda primavera fino all’inizio dell’autunno, protraendosi talvolta anche fino a ottobre inoltrato o inizi di novembre. -V. velutina forma colonie dalla vita abbastanza lunga, tracce di attività residua possono persistere fino a fine anno e qualche volta persino dentro gennaio.
Foto in alto: nido di Dolichovespula media dalla Repubblica Ceca Foto in basso: Nido di Vespa velutina ‘nigrithorax’ dalla Francia
We use cookies on our website to give you the most relevant experience by remembering your preferences and repeat visits. By clicking “Accept”, you consent to the use of ALL the cookies.
This website uses cookies to improve your experience while you navigate through the website. Out of these cookies, the cookies that are categorized as necessary are stored on your browser as they are essential for the working of basic functionalities of the website. We also use third-party cookies that help us analyze and understand how you use this website. These cookies will be stored in your browser only with your consent. You also have the option to opt-out of these cookies. But opting out of some of these cookies may have an effect on your browsing experience.
Necessary cookies are absolutely essential for the website to function properly. This category only includes cookies that ensures basic functionalities and security features of the website. These cookies do not store any personal information.
Any cookies that may not be particularly necessary for the website to function and is used specifically to collect user personal data via analytics, ads, other embedded contents are termed as non-necessary cookies. It is mandatory to procure user consent prior to running these cookies on your website.